Nagy Brett Manifesztum 2.
A Custers-effektus és a Brettanomyces Anyagcsere-dinamikája
Mivel sokan jeleztétek, hogy kemény a Brettes élet, így újra előrántjuk a hordó mélyéről azt, amitől még a legtapasztaltabb baktereknek is ketté áll a füle, a vad, ám zseni működésének az alapját, a Custers-hatást. Ízlelgessük: Custers, Custers.

Emlékeztetőként: a Brettanomyces, ez a kis szemétláda akkor is talál magának zabálnivalót a sörödben, amikor a rendes élesztők már rég harakirit követtek el. Ez a dög olyan, mint az a haverod, aki buli után még a hűtő mögé beesett száraz pizzaszélet is benyomja, majd feltőri a lelakatolt hűtődet is, de csak akkor, ha talál egy elektron-akceptort. Na de mi irányítja a kis bélpoklos szarzsák működését és ez miért, és legfőképpen hogyan befolyásolja a söröd ízvilágát (tudod a Custers-effektus). Meg különben is, ki volt Custers?
De előtte, a random kínzó kérdés, miért érdekes ez számunkra? Mert ennek mentén fogunk pár cikkel később mikró-oxidációt (MOX) számolni (lelki szemeim előtt látom, hogy itt már nem a következő imperial stout pattan fel, hanem egy vizespohárnyi brettes sörpárlat), ééés, ez alapján fogjuk górcső alá venni a házisörfőző berkekben használatos gyakorlati MOX megoldásokat. De mi az a górcső? Na ki tudja google nélkül? Szervesen kapcsolódik a témához.
Szóval Sir Henry Górcső, ja, ja nem, Custers. Custersről fogok értekezni. A hollandus bakterioloogus (1906.12.22. Swalmen, Hollandia) nélkül a "vad sör”(Michelle Wild után szabadon) kifejezés csak annyit jelentene, hogy valaki véletlenül beleejt egy marék lóhoz dörzsölt egeret a cefrébe, amit lássunk be, hogy nem annyira közelíti meg a hidegkómlózás ízvilágát. Ezért most ismerkedjünk meg Mathieu Theodoor Jozef Custersszel, a sörfőzés történetének egyik legvagányabb "élesztő-idomárjával".

Tudod, a legtöbb sörfőző élesztő (a Saccharomyces) olyan, mint a jóravaló stréber diák, aki időben lefekszik, csak azt eszi meg, amit elé raknak, és ha elfogy a kaja, szépen elalszik. De aztán ott van a Brettanomyces (vagy, ahogy mi hívjuk: - a Brett), ez a bőrdzsekis, láncdohányos punk, aki nem csak a hűtőt eszi ki, de ha nagyon éhes, még a konyhabútort is elrágcsálja miközben olyan szagot áraszt, mint az izzadt lószerszám. Mármint érted: zabla, kantár, nyereg, stb, nem az amire te gondoltál.
Sokáig senki nem értette meg, hogy miért csinálják ezt ezek a kis dögök, miért van az, hogy egyes esetekben kellemes trópusi ízeket, míg más alkalmakor fent részletezett bukét hozzák ki a sörből. Aztán jött Custers, ez a holland mikrobiolóógus zseni, kezében egy palack angol stock aleel és megfázott, bedugult orrhangon azt mondta: "Hé, skaacok, én rájöttem, miért ilyen fura ez a banda és mit miért csinál!" Custers a híres Technische Hoogeschool te Delft egyetemen tanított és kísérletezett, majd 1940-ben két spangli között letett az asztalra egy olyan doktori disszertációt, ami alapjaiban rázta meg a sörfőzést. Ő volt az első, aki módszeresen, tudományos alapossággal leírta és rendszerezte a Brettanomyces nemzetséget („Onderzoekingen over het gistgeslacht Brettanomyces” művében), valamint megfejtette a kis pöcsöknek a működésüket.[https://www.sciencedirect.com/topics/immunology-and-microbiology/brettanomyces]

Előtte is tudták (legalább is tudományosan 1904. óta), hogy léteznek ezek a "brit gombák" (innen a név: Brettano - brit, myces - gomba), de Custers volt az, aki rámutatott, hogy ezek nem csak véletlen szennyeződések, a sörfőzdék rémei, a Britt Izzadt Lószerszám Nemzeti Társaságának emblematikus kabbalái, hanem egy teljesen különálló, sajátos szabályok szerint játszó élesztők törzse. Ő jött rá, hogy miért képesek ezek a kis rohadékok túlélni és sörszerű italt varázsolni ott is, ahol a rendes élesztő már rég beadta a kulcsot.
Custers nélkül ma nem értenénk, hogyan kell kezelni egy Lambicot vagy egy vadale-t, érted: vad élt, persze ettől még korábban is elkészítették ezeket a söröket, de mos tmár tudjuk, hogy mi történik közben és hogyan szabályozzuk. Ő tanította meg nekünk, hogy a Brett nem csak erjeszt, hanem vadul bulizik a tartályban, aztán amikor elfogy a kaja, neki esik a tartály falának is, végül elájul és leülepszik az aljára, de közben olyan komplexitást hagy maga után, amitől eldobod az agyad. Custerst szokás idézni úgy, mint aki az egyik legkorábbi, rendszerezett Brett-kutatást végezte. A későbbi összefoglalók szerint több (egyes források szerint 17) különböző Brettanomyces-törzset írt le és jellemzett. Te meg nem tudsz felsorolni ebből ötöt sem. Nyugi, eddig én sem.[https://www.sciencedirect.com/topics/immunology-and-microbiology/brettanomyces]
A róla elnevezett jelenség a „Custers-effektus” (más néven negatív Pasteur-effektus) – Brettnél tipikusan azt a furcsa viselkedést jelöli, hogy szigorúan anaerob körülmények között az alkoholos fermentáció leállhat / hosszú lag fázisba mehet, de hidrogén-akceptorok (pl. kis oxigén, acetoin) jelenlétében ez a gátlás rövidülhet vagy megszűnhet és újra beindulhat az erjesztés. Persze ez nem minden Brett-törzsben azonosan jelentkezik, és gyakran átmeneti jellegű pl. klasszikus kísérletekben 7–8 óra után újra indulhat anaerob adaptációval.
Szóval, ha legközelebb egy fura, szaunázó ló bukéjú, füstős-fűszeres, ám de zseniális sört iszol, vagy éppen szembejön a palack matricán a Brettanomyces custersianus felirat, emeld meg a poharad Mathieu Theodoor Jozef Custersre. Szakirodalomban kifejezetten úgy szerepel, hogy ez utóbbi Brett Custers tiszteletére lett így elnevezve.

És akkor most csapjunk a szegfűborsba hempergett izzadt lovak közé, avagy hogyan üzemel a Brett. Na, ez tényleg fájni fog.
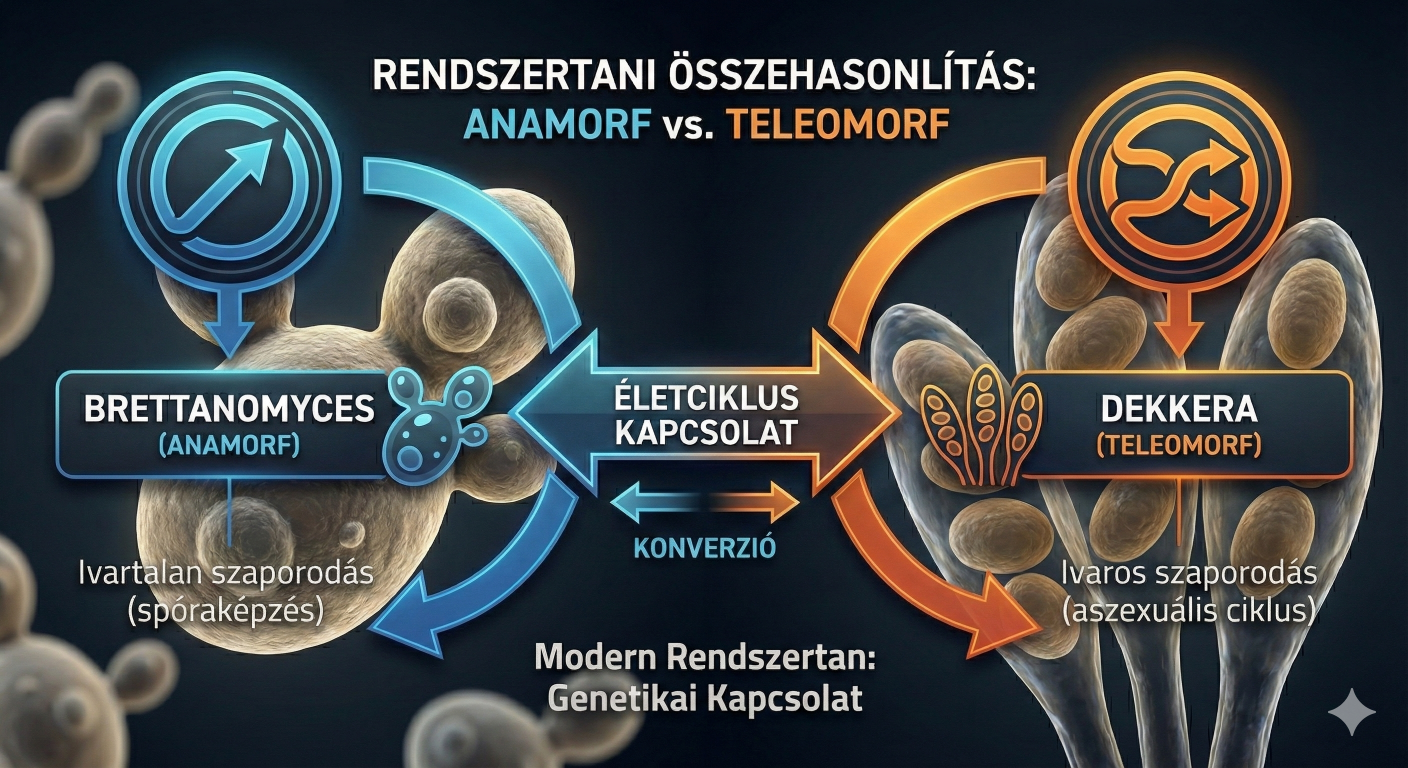
A sörfőzés mikrobiológiájának egyik legkomplexebb, ugyanakkor leginkább félreértett területe a Brettanomyces (teleomorf, ivaros alakban Dekkera) nemzetséghez tartozó élesztőgombák anyagcseréjének vizsgálata. Míg a konvencionális sörélesztők, elsősorban a Saccharomyces cerevisiae és Saccharomyces pastorianus metabolizmusa az évszázados ipari alkalmazásnak köszönhetően rendkívül jól feltérképezett, addig a különc Brettanomyces fajok élettani működése – különös tekintettel a redox-egyensúly fenntartására – számos rejtélyt és technológiai kihívást tartogat.
Ezen kihívások központi eleme a Custers-hatás, amelyet a szakirodalom gyakran „negatív Pasteur-effektusként” emleget, utalva arra a jelenségre, miszerint ezen organizmusok alkoholos erjedése anaerob körülmények között gátolt, míg oxigén jelenlétében stimulált. És hogy igazán fájjon, megpróbálom kellően kimerítő (engem legalább is az írás kimerített) részletességgel bemutatni a jelenség biokémiai hátterét, evolúciós okait, és ami a legfontosabb gyakorlati implikációit (hmm, micsoda kifejezés) a modern házisörfőzés számára.
Ebben a bejegyzésben nem csupán a Custers-hatás molekuláris mechanizmusait gyűjtöm össze és stresszellek bennetek, hanem azokat a szélesebb körű sörfőzési technológiákba is betekintünk (mint pl. a Brett starterkészítés, a hordós érlelés, palackozás és még a komlózási érdekességek is, stb. nyomokban megjelennek – mondjuk ezek már átcsúsznak a következő részbe).
És ha ez még nem rémísztett el eléggé, akkor végső kegyelemdöfésként röpke figyelmet szentelek a nitrát-asszimiláció nemrégiben feltárt szerepének, amely alapjaiban írja felül a korábbi ismereteket a Brett anaerob viselkedéséről.
A Brettanomyces nemzetség taxonómiai története önmagában is tükrözi a tudományos megértés fejlődését. Az N. Hjelte Claussen által 1904-es első leírás óta (később róla is megemlékszünk), amely a brit "stock ale"-ek másodlagos erjedéséért felelős organizmusként azonosította, a nemzetség megítélése a veszélyes romlási faktortól a nagyra becsült, komplexitást adó "terroir" elemig ingadozott. Ezen kettősség megértéséhez elengedhetetlen a Custers-hatás mélyreható ismerete, mivel ez a mechanizmus magyarázza a Brettek viselkedésének látszólagos kiszámíthatatlanságát, azaz a hosszú lappangási időket, a hirtelen ecetesedést, vagy éppen a váratlanul újra induló erjedést a palackban.
A Custers-hatás Biokémiai és Molekuláris Mechanizmusa
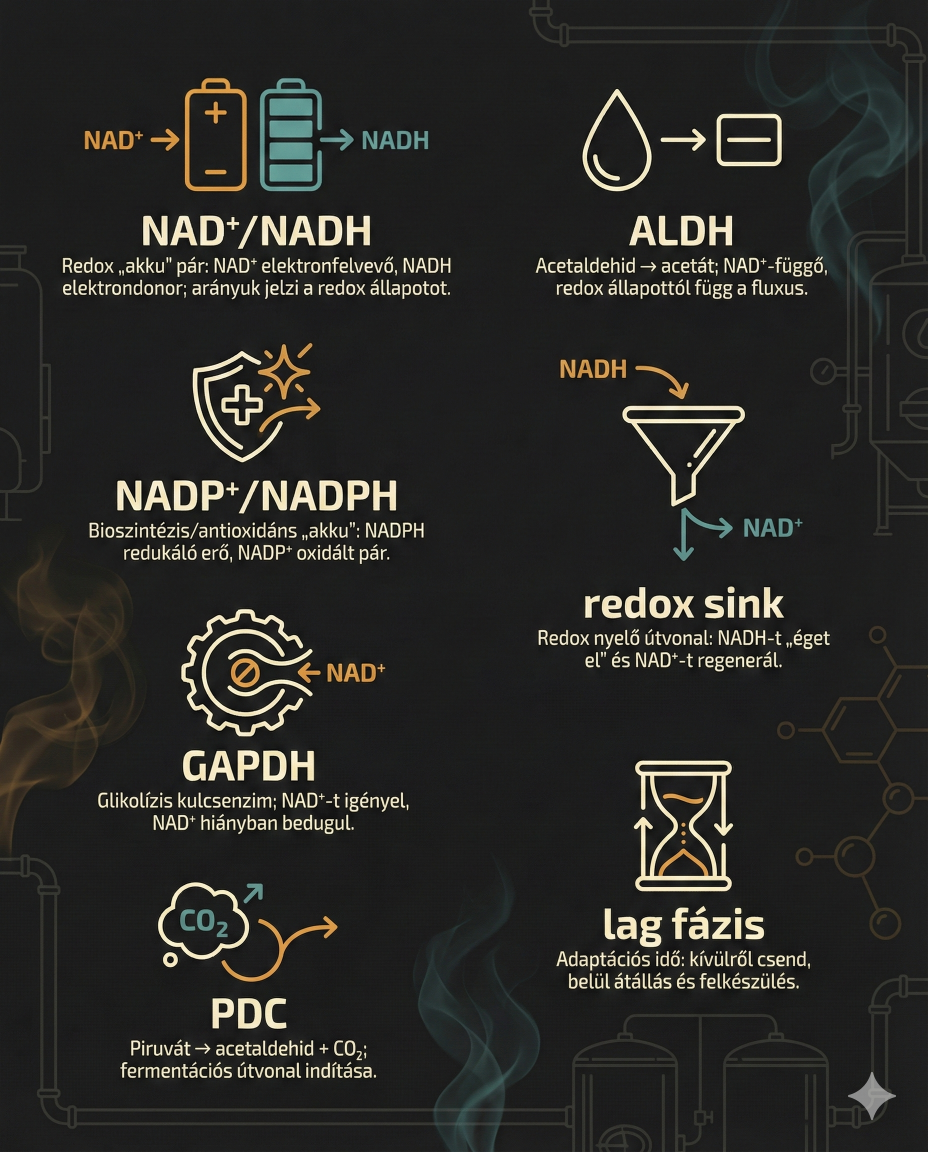
A Custers-hatás lényege egy alapvető metabolikus blokk, amely akkor lép fel, amikor a Brettanomyces sejtek hirtelen oxigénmentes (anaerob) környezetbe kerülnek. A jelenség megértéséhez a sejtszintű redox-folyamatok, különösen a nikotinamid-adenin-dinukleotid (NAD+/NADH) koenzim-rendszer egyensúlyának vizsgálata szükséges…
A NAD+/NADH Redox-egyensúly Központi Szerepe
Én szóltam, hogy fájni fog. Minden élő sejtben a glikolízis –azaz a cukrok lebontásának alapvető folyamata – során a glükózmolekula (cukor) oxidációja történik meg piruváttá ATP termelésével, de ez a folyamat egyúttal NAD+ redukcióját is igényli NADH-vá. A reakció kulcslépése a gliceraldehid-3-foszfát-dehidrogenáz (GAPDH) enzim által katalizált reakció. [https://pmc.ncbi.nlm.nih.gov/articles/PMC4869616/]
Gliceraldehid-3-foszfát + NAD⁺ + Pi → 1,3-biszfoszfoglicerát + NADH + H⁺ (GAPDH reakció)
Nézzük sorra a fenti káromkodást, avagy egy kis fogalommagyarázat. Szisszenjen az a brett ipa.
Glikolízis (nem, nem gyíkom Lizi, az más), alatt azt értjük, mikor a szőlőcukor (azaz a glükózmolekula) elbontodik piruvátmolekulává, miközben rengeteg energia, jelen esetünkben ATP és NADH keletkezik.
A piruvát, vagy vagy piroszőlősav anionja, az élő sejtek anyagcseréjének egyik legfontosabb kereszteződési pontja. Ez az a molekula, ahol a szénhidrátok, aminosavak és zsírok lebontása találkozik, és itt dől el, hogy a sejt mennyi energiát tud kinyerni az üzemanyagból. Kémiai képlete: CH3-CO-COO-. A piruvát a glikolízis végterméke. A folyamat során egy 6 szénatomos glükóz molekula két 3 szénatomos piruváttá bomlik le a citoplazmában.
ATP az az adenozin-trifoszfátot jelenti. Az ATP a sejtek elsődleges energiaforrása, a szervezet "biokémiai üzemanyaga", amely adeninből, ribózból (ez is cukormolekula) és három foszfátcsoportból áll. A rendszer a foszfátkötéseiben (tulajdonképpen ez az üzemanyag, mármint a foszfátkötés) raktározódó energiát szabadítja fel a sejtfolyamatokhoz, mint pl. az izommunka, a jelátvitel és bioszintézis, stb.
A NADH (nikotinamid-adenin-dinukleotid-hidrid) egy kulcsfontosságú koenzim, amely a sejtek energia-anyagcseréjében játszik központi szerepet azáltal, hogy elektronokat szállít az ATP termeléséhez, ezen kívül támogatja az agyműködést, javítja a mentális éberséget, és még antioxidáns tulajdonságokkal is rendelkezik. Ez a B3-vitamin (niacin) származéka, amely stabilizált formában étrend-kiegészítőként is kapható. Szerepe az energia-termelés: Elektronokat szállít a glükóz és zsírok lebontásakor, ami elengedhetetlen az ATP szintéziséhez. És mindezen felül támogatja a kognitív funkciók működését pl. serkenti az olyan neurotranszmitterek (pl. szerotonin, dopamin) termelését, amelyek javítják a hangulatot, a memóriát, a koncentrációt és az éberséget.
NADPH (Nikotinamid-adenin-dinukleotid-foszfát) a felépítő folyamatok (anabolizmus) és a védekezés fő szereplője. Fő feladata az „Építőanyag” biztosítása. Olyan folyamatokhoz kell, ahol a sejt új molekulákat gyárt (pl. zsírsavszintézis, DNS-másolás, stb.). Védelmi szerepkörben kulcsfontosságú az antioxidáns védekezésben (pl. a glutation regenerálásában) és segít semlegesíteni a káros szabadgyököket.
A sejt igyekszik az NADPH szintet magasan tartani (az NADP⁺-hoz képest), hogy mindig készen álljon a szintézisre.
GAPDH enzim (glicerinaldehid-3-foszfát-dehidrogenáz) egy kulcsfontosságú enzim, amely a glikolízisben, a glükóz energiává alakításának folyamatában vesz részt, de ezen kívül számos más funkciója is van, mint például a DNS-javítás, a transzkripció szabályozása, a sejtmagi transzport, az apoptozis (programozott sejthalál) és szerepet kap a neurodegeneratív betegségekben, például az Alzheimer-kórban.
A koenzim-A (CoA) egy koenzim, amely a zsírsavszintézis, a zsírsavoxidáció és a piruvát oxidáció (citromsavciklus) folyamataiban vesz részt. A fehérjék könnyen leválasztható része, egy nem-fehérje enzimalkotó; vagyis koenzim.
Na, most hogy ezek után a szomszéd borbély beretvapengéjét élezed egy izzadt lóbőrszíjon, megnyugodhatsz, tudod a legfontosabb alapokat. Igazából ezek csak nevek, hívhatnánk őket Katának, Viktornak, Tamásnak, Krisztiánnak, vagy éppen Ferinek is. Igazából a szerepük fontos.
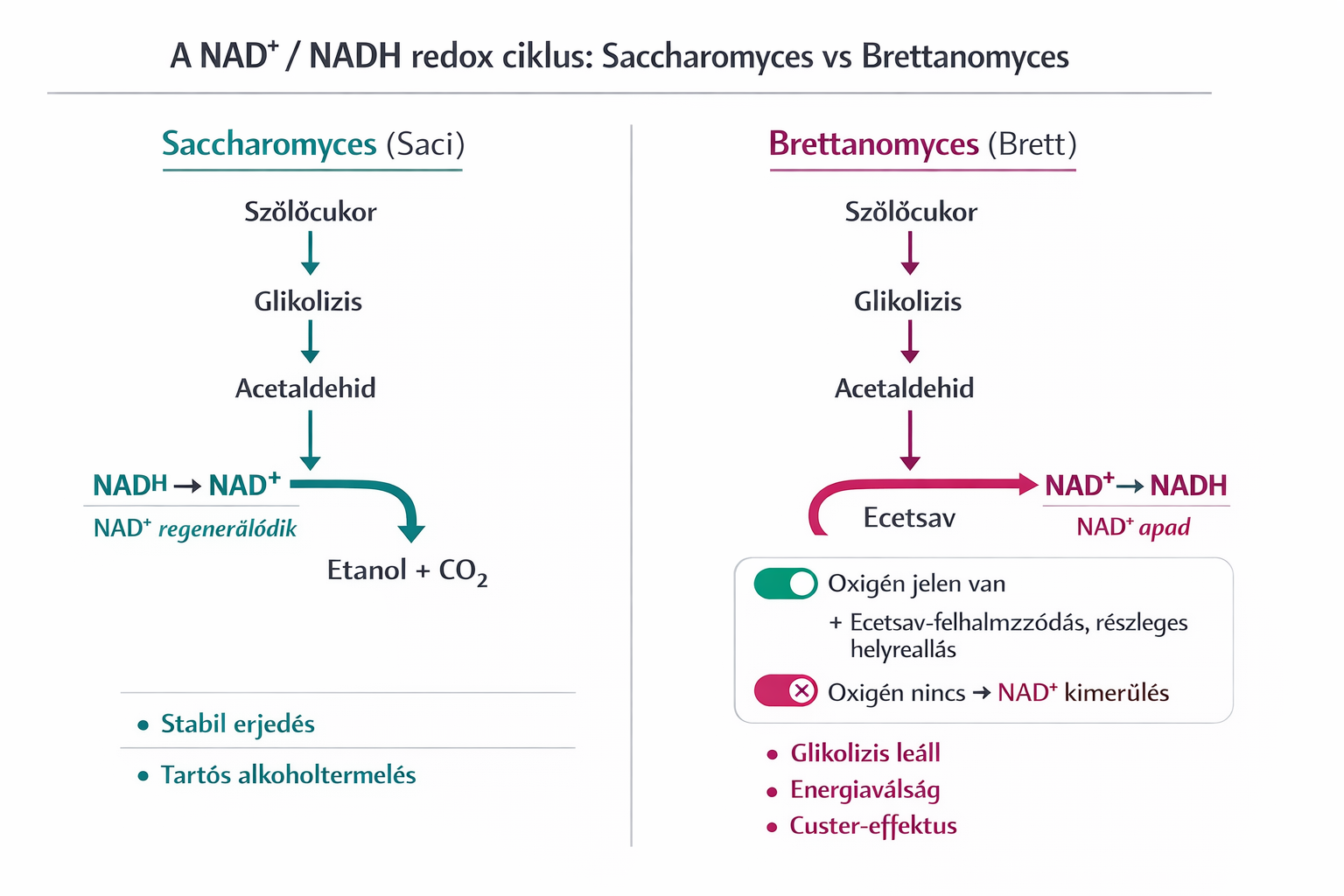
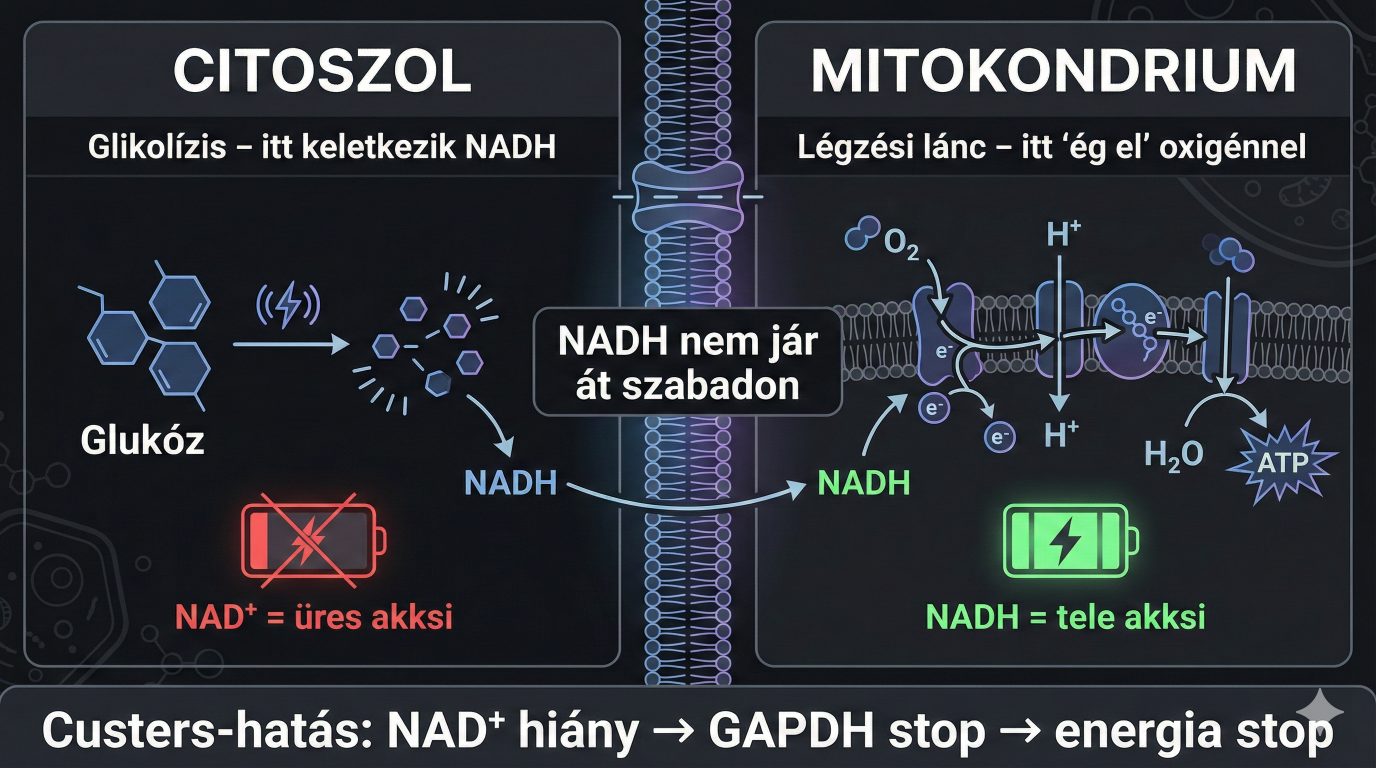
Alapozás után térjünk vissza a könnyed témánkhoz, tehát ahhoz, hogy a glikolízis folyamatosan termelhesse az energiát (ATP), a keletkező NADH-t folyamatosan vissza kell oxidálni NAD+-á. Ha ez a visszaoxidálás nem történik meg, a sejtben elfogy a NAD+, a GAPDH enzim tápanyag hiányában leáll, és ezzel együtt a teljes energiatermelő anyagcsere összeomlik. A Saccharomyces cerevisiae (Sacci a sörélesztő) és más sejtek esetében, mivel a NADH nem képes átjutni a mitokondriális membránon, külön mechanizmusok oxidálják a citoszolikus és a mitokondriális NADH-t. Anaerob körülmények között, ahol az oxidatív foszforiláció (légzés) nem működik, a Saccharomyces két fő útvonalat használ a citoszolikus NADH visszaoxidálására ("redox sink")
- Alkoholos erjedés: Az acetaldehid redukciója etanollá az alkohol-dehidrogenáz (ALDH) által. Ez a lépés regenerálja a glikolízis során felhasznált NAD+-t.
- Glicerin termelés: Amikor a biomassza-képződés vagy egyéb bioszintetikus folyamatok miatt többlet NADH keletkezik, a Saccharomyces dihidroxi-aceton-foszfátot (DHAP) redukál glicerin-3-foszfáttá, majd glicerinné. Ez a folyamat extra NAD+-t generál, fenntartva a redox-egyensúlyt.
Most, hogy jó sokat káromkodtam, ezzel már tudjuk, hogy mit jelent a NADH (ugye a tele akksi) de nem tudjuk mit jelent a citoszolikus és a mitokondriális NADH. Az anyagcsere-folyamatok megértéséhez kulcsfontosságú a NADH sejten belüli elhelyezkedése, mivel a sejt különböző részei (kompartmentjei) között ez a molekula nem tud szabadon átjárni. A citoszol a sejt "alapállománya", ahol az sejtszervecskék úsznak. A citoszolikus NADH az erjedés során a sejtfolyadékban keletkező "hulladék", amit újra kell hasznosítani (NAD+-á alakítani), hogy a cukorbontás menjen tovább. A mitokondriális NADH pedig a sejt belső égetőművében lévő üzemanyag, ami csak oxigén jelenlétében tud hatékonyan hasznosulni, viszont itt a hangsúly a hatékonyan van, mivel oxigén nélkül is képes hasznosulni, de nagyon rossz hatásfokkal. A Custers-hatás alatt ez a két raktár közötti egyensúly felbomlik, leegyszerűsítve.
A Glicerin-blokk és a Brettanomyces anyagcsere-csapdája
A Brettek, különösen a B. bruxellensis és B. intermedius (a modern taxonómia szerint gyakran szinonimák vagy közeli rokonok) esetében ez a rendszer alapvetően hibás. A kutatási adatok egyértelműen mutatják, hogy ezek az élesztők anaerob körülmények között képtelenek vagy csak elhanyagolható mértékben képesek glicerint termelni. Ennek a hiányosságnak a molekuláris oka valószínűleg a glicerin-3-foszfát-foszfatáz (GPP) enzim aktivitásának hiánya vagy annak rendkívül alacsony szintje. Saccival ellentétben, amely a GPD1 és GPD2 gének expressziójával aktívan szabályozza a glicerintermelést a redox-egyensúly fenntartása érdekében, a Brettanomyces nem rendelkezik ezzel a biztonsági szeleppel. [https://www.researchgate.net/figure/The-Custers-effect-of-B-D-bruxellensis-A-Schematic-overview-of-main-factors_fig1_275587397]
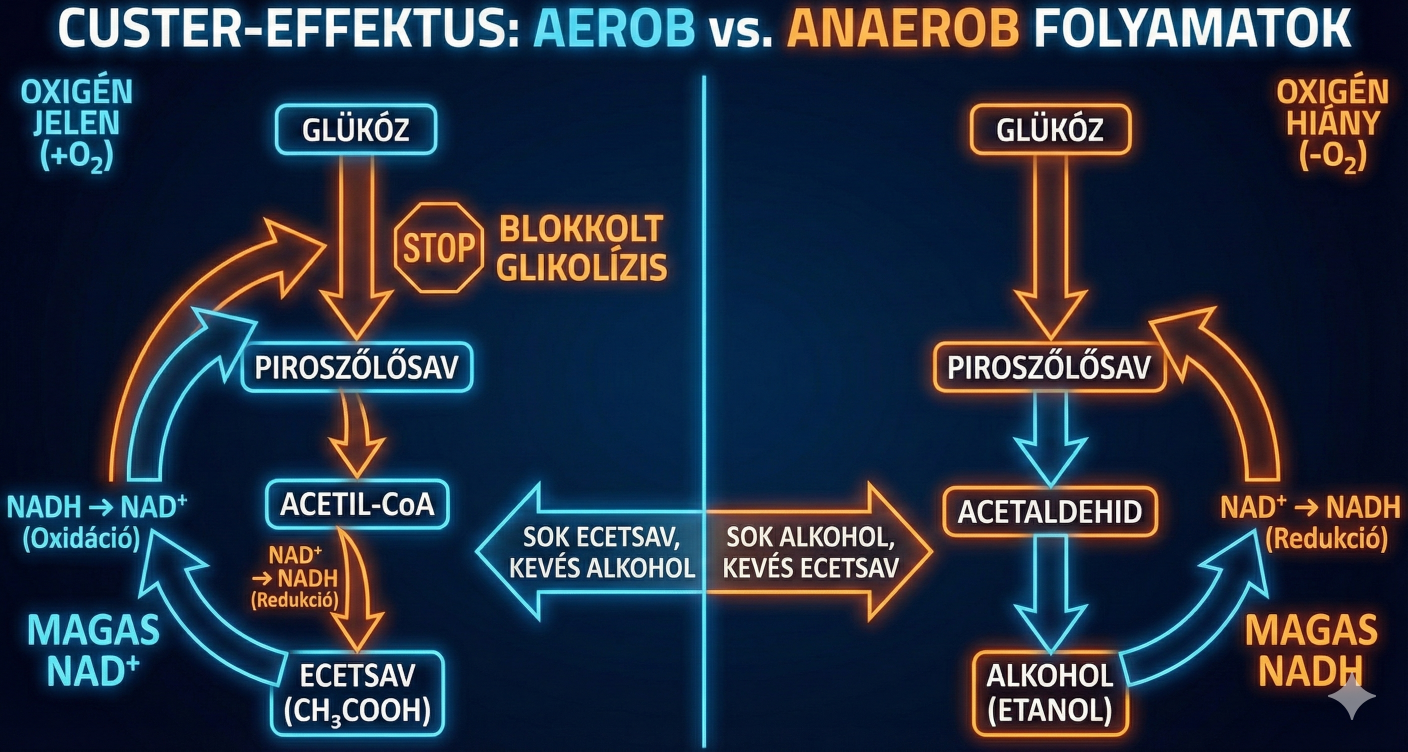
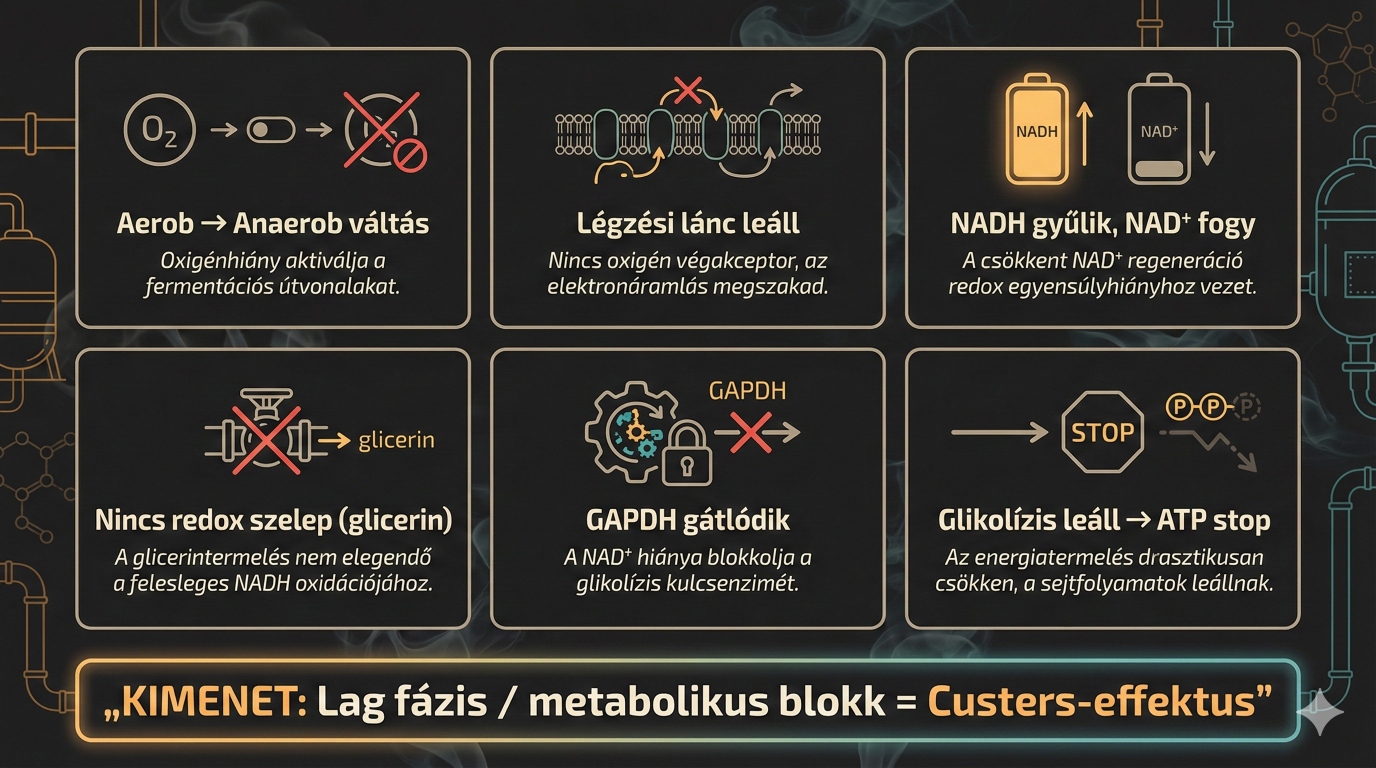
Amikor egy Brett kultúrát aerob környezetből hirtelen anaerob környezetbe helyezünk (például palackozáskor vagy egy zárt erjesztőtartályba való átfejtéskor), a következő katasztrófa-szekvencia (imádom ezt a fogalmat) játszódik le a sejtben:
- Az oxigén, mint végső elektronakceptor eltűnik, így a mitokondriális légzési lánc leáll.
/*A mitokondriális légzési lánc kifejezés a biokémia egyik legfontosabb folyamatára, a sejtlégzésre utal. Hogy érthető legyen, képzelj el egy "elektron-stafétát", ahol a sejt a cukorból kinyert energiát (elektronokat) adogatja kézről kézre. Amikor tehát azt hallod, hogy "az oxigén, mint végső elektronakceptor eltűnik", gondolj arra, hogy elfogyott az egyetlen "elektron-staféta" molekula, amely képes lett volna biztonságosan továbbítani/felvenni a sejtben felhalmozódó elektronokat.*/
- A glikolízis folytatódik, NADH-t termelve.
- A biomassza-építés és egyéb mellékreakciók további NADH-t generálnak.
- Glicerin-termelés hiányában nincs alternatív útvonal a felesleges NADH visszaoxidálására.
- Az alkoholos erjedés önmagában redox-semleges (1 NADH keletkezik, 1 NADH fogy), így nem képes eltüntetni a bioszintézisből származó felesleget.
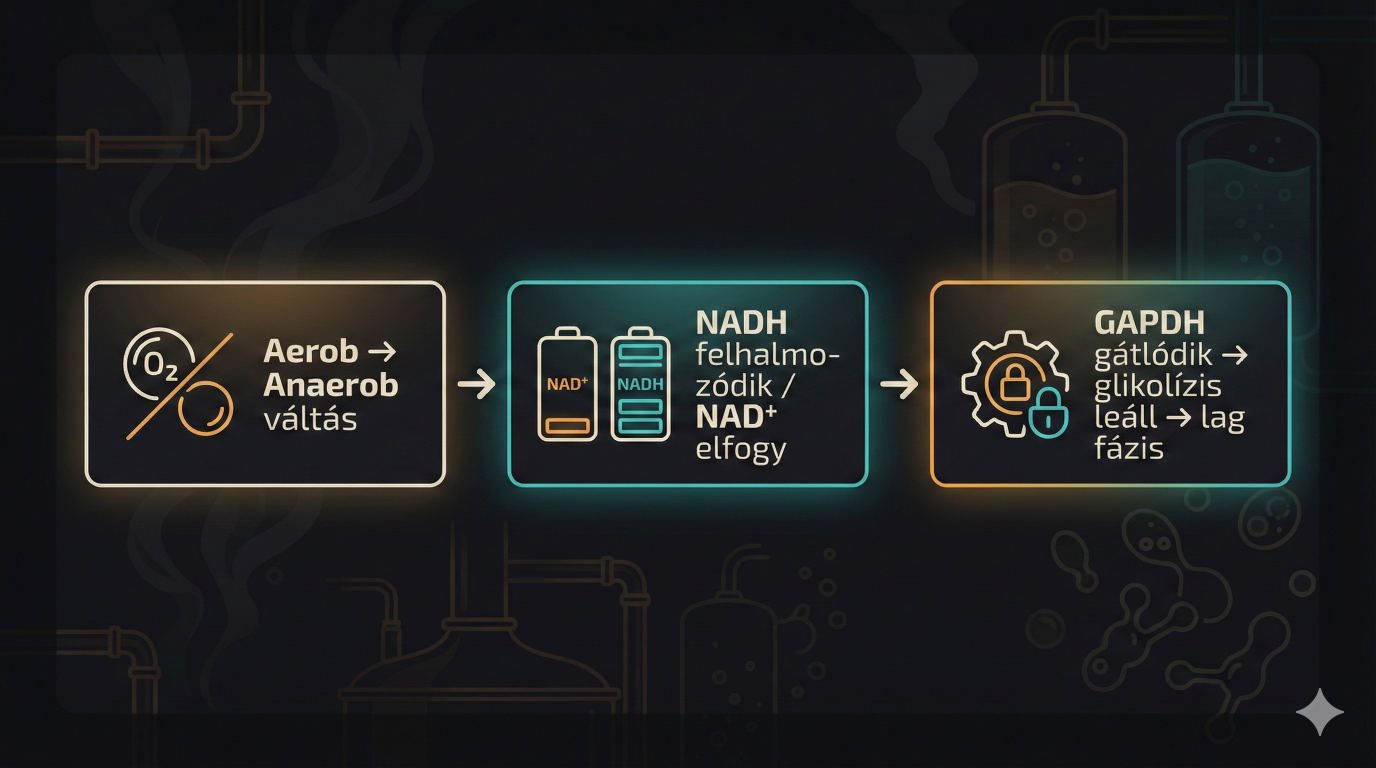
- A NAD+/NADH arány drasztikusan lecsökken (a NADH felhalmozódik).
- NAD+ hiányában a GAPDH enzim gátlódik.
- A glikolízis leáll, az ATP termelés megszűnik, és a sejt egyfajta "hibernált", nem növekvő állapotba, az úgynevezett Lag fázisba kerül.
Ez a metabolikus blokk maga a Custers-effektus. A "lag fázis" hossza változó lehet, tarthat néhány órától akár hetekig vagy hónapokig is, attól függően, hogy a sejt talál-e alternatív elektronakceptort, vagy lassan, "szivárgó" anyagcsere-utakon keresztül képes-e adaptálódni. A sörfőzés biokémiájában ezek a kifejezések a Brett túlélési stratégiáit jelentik. Amikor az elsődleges "autópálya" (az oxigénes légzési lánc) lezárul, az élesztő kénytelen "mellékutakat" vagy "B-terveket" keresni, hogy ne fulladjon bele a saját NADH-felhalmozódásába. És pontosan ez az (a spóraképzésen felül) amely brutál ellenállóvá teszi a kis különc dögöt, a láncdohányos, ananászos pizzát zabáló, vörösboroskólát szürcsölő punk-rock-soul-beat-heavyjazz-metalt hallgató, mangópálinkába áztatott sajtosstanglival öblögető barátunkat.
Mit jelent ez a gyakorlatban?
- Alternatív elektronakceptorok:
- Képzeld el, hogy a sejtnek "szemetet" (elektronokat) kell ürítenie. Az oxigén a nagy szemétszállító kamion. Ha az oxigén nincs ott, a sejt keres másokat, akik elvisznek egy-egy zacskót.
- Ilyen "alkalmi fuvarosok" a nitrátok (amik sok gyümölcsben benne vannak). Ezért van az, hogy egy megrekedt Brett-fermentáció leggyakrabban beindul (nem minden esetben!), ha meggyet vagy málnát (gyümölcsöt) dobsz a sörbe a gyümölcsben lévő nitrátok átveszik az elektronokat, felszabadul a NAD+, és a motor újraindul. Ez lesz a fent említett nitrát-asszimiláció.
- Ez az Brett élesztő "túlélő üzemmódja". Ha nincs se oxigén, se gyümölcs, se nitrát a kútvízből, a sejt rendkívül lassan, nem hatékony mellékutakon keresztül (pl. aminosavak gyártása vagy nagyon lassú észter termelés közben) kezd el megszabadulni az elektronoktól.
- Ez nem egy "tervezett" főút, inkább olyan, mintha a fal repedésein szivárogna át a víz. Nagyon kevés NAD+ regenerálódik, de hosszú hetek vagy hónapok alatt ez a kevés is elég ahhoz, hogy a sejt végül lassú növekedésnek induljon, és mindenféle aromát termelhessen, vagy éppen nem. Nem szükségszerű az aromatermelés.
Összegezve: A Brett nem feltétlenül áll le örökre oxigén nélkül. Vagy talál egy másik molekulát, ami segít neki (akceptor), vagy a saját belső, lassú és nem hatékony folyamatait használva "kivárja", amíg elég energiát gyűjt a folytatáshoz. Vagy megdöglik, de ez a legritkább változat.
/*A fenti felsorolásban az egyes(1) pontban olvashatsz egy „a mitokondriális légzési láncról” ez (más néven elektronszállító lánc) a sejt "erőművének", a mitokondriumnak a legbelső részén található biokémiai gépezet. Ez a lánc felelős azért, hogy a tápanyagokból (cukorból) kinyert energiát a sejt számára használható "üzemanyaggá" (ATP-vé) alakítsa.*/
Az Oxigén és az Ecetsav Paradoxona
A Custers-hatás feloldása oxigén bevezetésével történik. Ez első pillantásra logikusnak tűnik, mert az oxigén jelenlétében a légzési lánc működik, a NADH visszaoxidálódik, és a NAD+ készlet helyreáll. Hát kurvára nem. A Brett reakciója az oxigénre sajátos és a sörfőzők számára gyakran problémás. Aerob körülmények között a Brettanomyces nemcsak a légzést, hanem egy erőteljes, fermentatív jellegű anyagcserét is folytat, amelynek egyik fő terméke az ecetsav. Ez a folyamat az ún. "PDC-bypass" (Piruvát-DeKarboxiláz kerülőút) amely pontosabban az acetaldehid-dehidrogenáz útvonal felerősödését jelenti. A reakciósorozat a következő:
- Piruvát → acetaldehid + CO₂ (piruvát-dekarboxiláz, PDC)
- Acetaldehid + NAD(P)⁺ + H₂O → acetát (ecetsav) + NAD(P)H + H⁺ (aldehid-dehidrogenáz, ALDH)
Itt érhető tetten a Brettanomyces evolúciós stratégiájának paradoxona, azaz az ecetsav termelése további NADH-t termel. Anaerob körülmények között ez végzetes lenne, hiszen tovább súlyosbítaná a Custers-hatást. Azonban oxigén jelenlétében a sejt "megengedheti magának" ezt a pazarlást, mivel a légzési lánc képes elnyelni a keletkező többlet elektronokat (NADH-t), miközben az ecetsav termelése extra ATP-t vagy metabolikus előnyt biztosít a vetélytársakkal szemben (az ecetsav toxikus sok más mikrobára).
A Brettanomyces ALDH enzimjei közül a NAD+-függő forma glükóz jelenlétében inaktiválódik, míg a NADP+-függő forma aktív lehet. A kutatások azt mutatják, hogy a Brett az acetátot nem képes hatékonyan tovább bontani acetil-CoA-vá és a Krebs-ciklusba (Nyugi, élesztők működésénél lesz róla szó bőven) juttatni, vagy az acetil-CoA-szintetáz aktivitása nem elegendő ehhez, ezért az ecetsav felhalmozódik a sejtben, majd kiválasztódik a közegbe. [https://pubmed.ncbi.nlm.nih.gov/6115615/]
Ez a mechanizmus magyarázza, miért termel a Brett hatalmas mennyiségű ecetsavat fél-aerob (mikroaerofil) körülmények között. Ha a rendszerbe folyamatosan kis mennyiségű oxigén jut (pl. hordó dongáin keresztül, vagy rosszul záró erjesztőtartályban), az éppen elegendő a Custers-blokk feloldásához (a NADH visszaoxidálásához), de a sejt anyagcseréje az ecetsav-termelés irányába tolódik el, mint egyfajta "túlfolyó" mechanizmus. És ha ez túl van tolva, mert sokszor szelfizel a nyitott erjesztőddel, akkor kapod az igen hivalkodó ízű sörecetet, amitől még a vízkő is elmenekül a karsztbarlang nyilvános wécéjéből. [https://rex.libraries.wsu.edu/view/pdfCoverPage?instCode=01ALLIANCE_WSU&filePid=13406943260001842]

A "Negatív Pasteur-effektus" Terminológiai Tisztázása
Louis Pasteur eredeti megfigyelése (Pasteur-effektus) azt írta le, hogy oxigén jelenlétében az élesztők cukorfogyasztása és alkoholtermelése csökken, mivel a légzés sokkal hatékonyabb energiatermelést tesz lehetővé, mint az erjesztés (36 ATP/glükóz vs. 2 ATP/glükóz), így kevesebb cukorra van szükség. Ezzel szemben a Custers-hatásnál az oxigén serkenti az erjedést (az etanol és ecetsav termelését), míg az anaerob körülmények gátolják azt. Ezért nevezte el Scheffers "negatív Pasteur-hatásnak". Fontos megkülönböztetni ezt a Kluyver-effektustól (bizonyos cukrok erjesztésének képtelensége anaerob módon, de légzése lehetséges) és a Crabtree-effektustól (magas cukorkoncentráció esetén a légzés gátlása és erjedés preferálása még oxigén jelenlétében is). Rossz hírem van, ezekről is lesz szó későbbiekben az élesztők működésénél. A Brettanomyces általában Crabtree-negatívnak minősül bizonyos források szerint, bár ez törzsfüggő lehet, és viselkedése a Custers-hatás dominanciája miatt egyedi kategóriát képez. [https://www.geeksforgeeks.org/biology/pasteur-effect-notes/]
Az alábbi táblázat részletesen összehasonlítja a Saccharomyces és a Brettanomyces metabolikus válaszait az eltérő oxigénszintre.
A Pasteur-effektus és a Custers-effektus összehasonlítása
|
Jellemző |
Pasteur-effektus (Saccharomyces cerevisiae) |
Custers-effektus (Brettanomyces bruxellensis) |
|
Kiváltó környezeti tényező |
Oxigén bevezetése anaerob kultúrába. |
Oxigén elvonása aerob kultúrából. |
|
Metabolikus válasz |
A glikolízis és fermentáció lassulása, átállás légzésre. |
A glikolízis és fermentáció drasztikus lassulása vagy leállása (Lag fázis). |
|
Glicerin termelés |
Jelentős anaerob körülmények között (Redox szelep). |
Hiányzik vagy elhanyagolható anaerob körülmények között. |
|
Redox egyensúly (NAD+/NADH) |
Fenntartott glicerin és etanol útvonalakon keresztül. |
Felborul anaerob körülmények között (NADH felesleg). |
|
Ecetsav termelés |
Általában alacsony, stresszválaszként nőhet. |
Magas, különösen aerob/mikroaerofil körülmények között (ALDH aktivitás). |
|
Oxigén hatása a fermentációs rátára |
Gátolja (a légzés hatékonyabb ATP termelése miatt). |
Stimulálja (a redox blokk feloldása miatt). |
|
Evolúciós előny |
Energiahatékonyság maximalizálása. |
Túlélés változó oxigénszintű (pl. gyümölcshéj) környezetben, ecetsavval a versenytársak gátlása. |
Forrás: https://scispace.com/pdf/metabolism-of-acetaldehyde-and-custers-effect-in-the-yeast-1djtfkf4y8.pdf
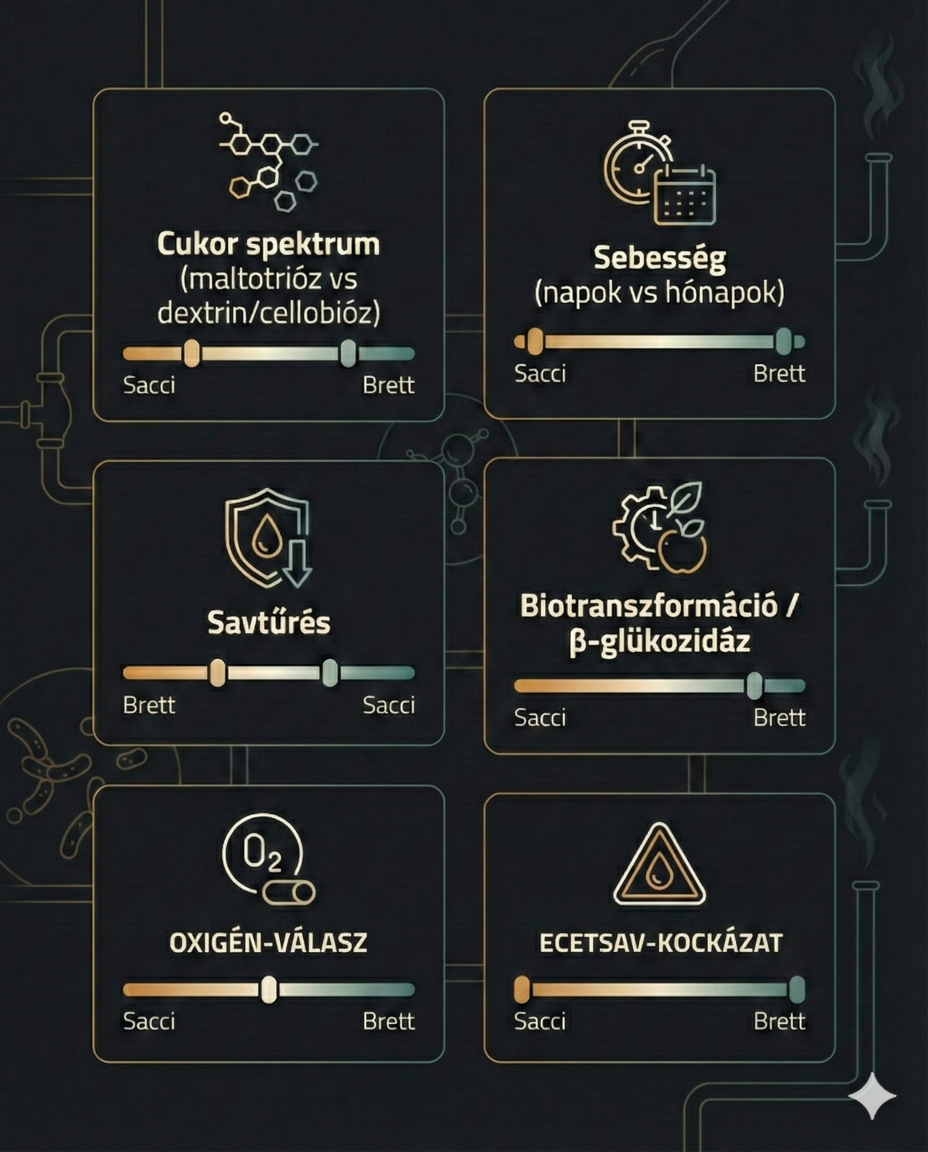
Brett vs. Saccharomyces (A Nagy Párbaj)
|
Tulajdonság |
Saccharomyces |
Brettanomyces |
|
Oxigén hatása |
Pasteur-effektus (Oxigén gátolja az erjedést) |
Custer-effektus (Oxigén serkenti az ecetgyártást, anaerob blokk) |
|
Cukorlebontás |
Csak egyszerű cukrok (maltotriózig) |
Mindent is (dextrinek, cellobióz) -> Szuper-attenuáció |
|
Béta-glükozidáz |
Ritka / Gyenge |
Erős (Komló illatfelszabadítás!) |
|
Spóraképzés |
Gyakori |
Ritka (csak Dekkera formában) |
|
Sebesség |
Gyors (napok) |
Lassú (hónapok) - kivéve 100% oltásnál |
|
Savtűrés |
Közepes |
Extrém (pH 3.0 alatt is elcsövezik) |
Tádádádádámmm: Nitrát Asszimiláció, A Custers-effektus "Achilles-sarka" és Megkerülése
A Brettanomyces kutatás egyik legizgalmasabb új fejleménye a nitrát-anyagcsere és a Custers-effektus közötti közvetlen kapcsolat feltárása. Míg Sacci a sörélesztő (S. cerevisiae) nem képes a nitrátot nitrogénforrásként hasznosítani, addig a Brettanomyces fajok többsége rendelkezik ezzel a képességgel, és ez alapvetően befolyásolja az anaerob viselkedésüket.
A Nitrát, mint Alternatív Elektronakceptor
A nitrát (NO₃⁻) asszimilációja során a sejteknek redukálniuk kell a nitrátot ammóniává (NH4+ hogy beépíthessék az aminosavakba és nukleotidokba. Ez a redukciós folyamat elektronokat igényel, amelyeket a NADH (vagy NADPH) szolgáltat.
A reakcióútvonal a következő enzimeket és géneket foglalja magában.
- Nitrát Transzporter (YNT1): A nitrát felvétele a sejtbe.
- Nitrát-reduktáz (YNR1): NO₃⁻ + NAD(P)H + H⁺ → NO₂⁻ + NAD(P)⁺ + H₂O
- Nitrit-reduktáz (YNI1): NO₂⁻ + 3 NAD(P)H + 5 H⁺ → NH₄⁺ + 3 NAD(P)⁺ + 2 H₂O
Látható, hogy egyetlen nitrát molekula ammóniává történő redukciója során 4 molekula NAD(P)H oxidálódik NAD(P)+-á. Ez a folyamat rendkívül hatékony "elektronnyelőként" működik. Anaerob körülmények között, ahol az oxigén hiányzik és a glicerin-útvonal nem működik, a nitrát jelenléte lehetővé teszi a Brettanomyces számára, hogy megszabaduljon a felesleges NADH-tól.
Ezzel fel lehet oldani a Custers-hatást, azaz újra be lehet durrantani az erjesztését, vagy marhára lecsökkenteni a lag-fázist.
Képletesen szólva képzeld el, hogy a Brett ebben az esetben olyan, mint egy tivornyázó banda a sörödben, akiknél hirtelen eldugult a klotyó, így ott állnak a sötétben, kezükben a piával (a cukorral), de nem merik lehúzni, mert kicsordulna. Na, ilyenkor jön a nitrát, ami gyakorlatilag egy ipari szippantóskocsi sebességével takarítja el az útból a szart, hogy mehessen tovább a buli. És mindez a sörödben, az erjesztőben.
Kísérleti bizonyítékok igazolják, hogy ha nitrátot adunk egy anaerob Brett tenyészethez, akkor a lag fázis drasztikusan lerövidül vagy teljesen eltűnik, és beindul az erjedés, vagy zavartalanul folyik tovább. Ez a mechanizmus gyakorlatilag "kikapcsolja" a Custers-hatást, mivel a nitrát helyettesíti az oxigén szerepét a redox-egyensúly helyreállításában. Ez a felfedezés forradalmi jelentőségű a sörfőzésben. Azt jelenti, hogy a sörlé nitráttartalma meghatározó tényező abban, hogy a Brettanomyces milyen gyorsan és milyen karakterrel erjeszt.[https://pmc.ncbi.nlm.nih.gov/articles/PMC4257070/]
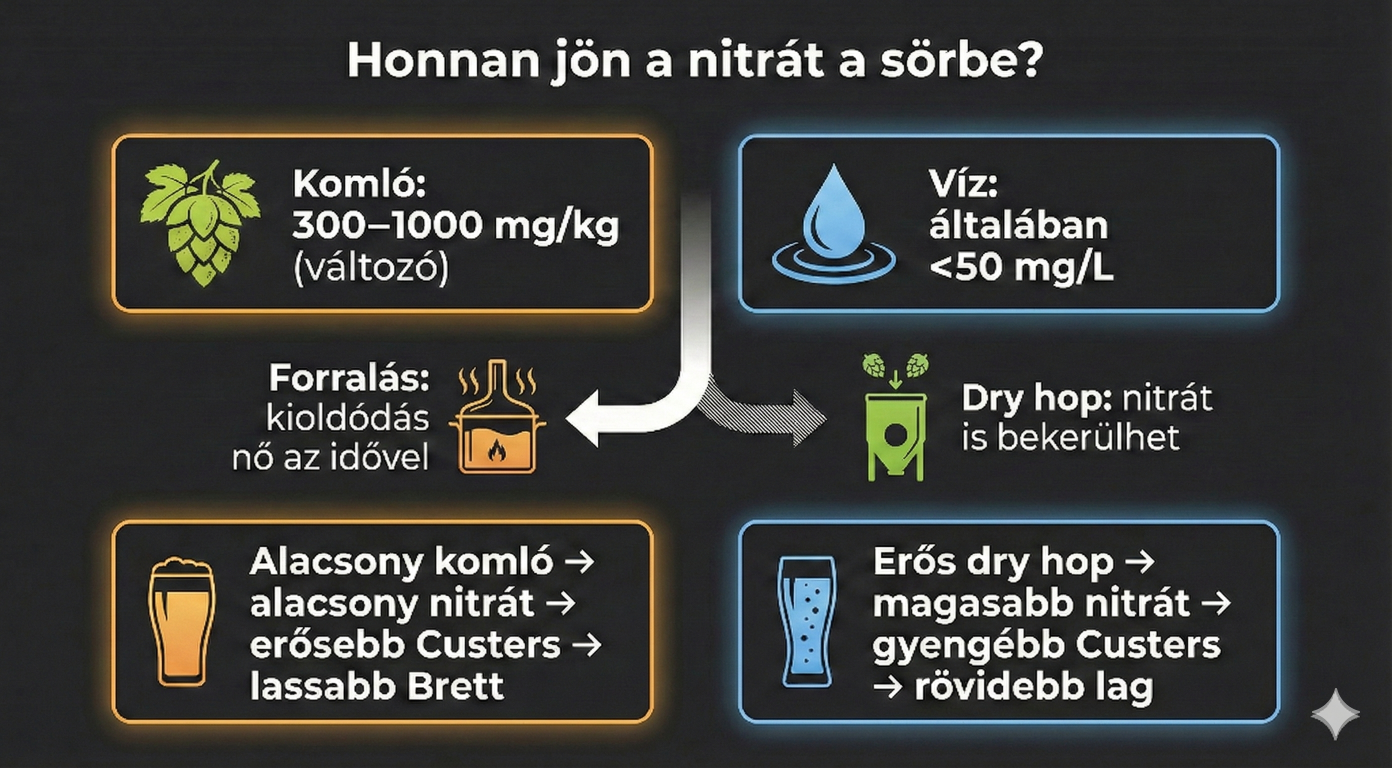
A Nitrát Forrásai a Sörfőzésben: Komló és Víz
Honnan kerül nitrát a sörbe? Nem, nem a rockbanda fogja belebulizni, hanem, ééés itt jön a meglepetés: a legjelentősebb forrás meglepő módon a komló. A komlók nitráttartalma fajtától és termesztési körülményektől függően széles skálán mozoghat (300–1000 mg/kg). A sörfőzés során a nitrátok kioldódása a forralási idővel lineárisan nő, de a hidegkomlózás (dry hopping) során is jelentős mennyiség kerülhet a sörbe. És való igaz az izzadt lóhoz dörgölt döglött egérben is sok a nitrát (drymice). Ez különösen releváns a modern, erősen komlózott sörstílusok (pl. NEIPA, modern Farmhouse Ale) esetében.
- Hagyományos, alacsony komlótartalmú sör: Alacsony nitrátszint >> Erős Custers-effektus >> Lassú Brett aktivitás, hosszú érlelés szükséges.
- Erősen hidegkomlózott sör: Magas nitrátszint >> Gyenge vagy megszűnő Custers-effektus >> Gyors Brett erjedés, rövidebb lag fázis.
A sörfőzővíz nitráttartalma is hozzájárulhat ehhez (bár az ivóvíz szabványok ezt korlátozzák, általában 50 mg/l alatt), így a komlókoncentráció a döntő tényező. A Brett képes már egészen alacsony, 6 mg/l nitrogénkoncentráció mellett is növekedni, azonban a Custers-effektus feloldásához szükséges (sztöchiometriai) mennyiség a redox-egyensúlytól függ. Ez nem jelenti azt, hogy fogod a konzervsót (kálium-nitrát), lőport, nagyfater salétromsav készletét és határozott mozdulattal telepumpálod a cefrédet ezekkel, mert az instant halál.
Brettek Genetikai Variabilitása a Nitrát Asszimilációban /BGVNA/
Fontos megjegyezni, hogy nem minden Brettanomyces törzs egyforma. A Carlsberg Kutatólaboratórium és más intézetek genomikai vizsgálatai kimutatták, hogy a nitrát-asszimilációs génklaszter (YNR, YNI, YNT) megléte vagy hiánya törzsspecifikus. [https://pmc.ncbi.nlm.nih.gov/articles/PMC4257070/]
- Nitrát-pozitív törzsek (pl. CRL-1): Rendelkeznek a megfelelő génklaszterrel, képesek hasznosítani a nitrátot, így gyorsabban erjesztenek komlózott sörlében anaerob körülmények között.
- Nitrát-negatív törzsek (pl. CRL-27): Hiányzik belőlük a specifikus génklaszter, vagy inaktív. Ezeknél a törzseknél a Custers-effektus sokkal erőteljesebben érvényesül, és a nitrát jelenléte nem segíti őket a lag fázis leküzdésében.
Ez a genetikai különbség magyarázhatja, miért viselkednek eltérően a különböző kereskedelmi Brett keverékek ugyanabban a sörlében. És erről majd később bővebben is vekengünk.
A Nagy Brett Manifesztum következő részében már tényleg beszélünk az ízképzésről és észterekről, valamint a komló biotranszformációról. A biokémia újabb, szagosabb tájékára evezünk és elmerülünk a menny és pokol alkémiájában, az észterek sűrűjében, megnézzük hogy milyen az, amikor a Brett gyümölcsnapot tart, és részt veszünk A Nagy Banán-Gyilkosságban (észteráz aktivitás). Az egészet megfűszerezzük Funky Fenolokkal közel merészkedve a Sörfőzők Rémálmához, az Egérízhez (Mousiness).
Ezt követően részben elmélkedünk brett starterek készítésről, oltási stratégiákról, palackbombák nemkészítéséről, Szuper-attenuációról éééés a hordós érlelésbe is belekóstolunk. Ez utóbbi fog megágyazni nekünk a mikró-oxidáció (MOX) gyakorlatba ültetésének a házisörfőzésben.
----------------------------
Hálásan köszönöm mindenkinek az eddigi támogatásokat, elindult az oldal migrációjának előkészítése és az új wordpress dizájn felépítése.
Támogatás
A Parabeer blog jelenleg ingyenes felületen fut, ezt nyilván látod, érzékeled.
Megcéloztam az átköltözést egy saját domainre, reklámmentes, hosszú távon fenntartható szakmai oldalt létrehozva így.
Ha értékesnek találtad a cikket,
meghívhatsz egy virtuális brettes sörre itt:
https://buymeacoffee.com/parabeer
Köszönöm – minden korty számít.